La genética es la ciencia que estudia la herencia biológica,
es decir, la transmisión de los caracteres de generación en generación.
Comprende todos los factores que influyen y que se transmiten de un pájaro a
sus descendientes, de todas las variedades y razas de canarios. Sólo considerar
que se aplica principalmente a la canaricultura de color, donde se cumplen
claramente las reglas. No tanto en canaricultura de postura, ya que en esas
razas sólo se tiene en cuenta el aspecto exterior, independientemente de que
sean machos o hembras. Tampoco en la canaricultura de canto, ya que sólo se
manifiesta en el macho y no se rige exclusivamente por el patrón genético, sino
que intervienen otros factores.

Gregor Mendel descubrió que los
genes se transmiten de una forma sorprendentemente sencilla. Hasta ese momento
nadie lo había observado con claridad porque no se habían parado a observar
características concretas de los seres. Mendel lo hizo, primero con guisantes,
después con flores y maíz.
En la transmisión genética, las células se pueden dividir mediante dos
modalidades: Mitosis y meiosis. En la mitosis se obtienen dos células hijas con
el mismo número de cromosomas que la célula madre e idénticas entre sí. En la
meiosis se obtienen cuatro células hijas con la mitad de cromosomas que la
célula madre y genéticamente diferentes. La división meiótica tiene lugar
durante los procesos de gametogénesis (espermatogénesis y ovogénesis), es
decir, durante la formación de los gametos. Mediante la meiosis se mantiene
constante el número de cromosomas en los individuos de la misma especie, ya que
se separan los pares de cromosomas homólogos.
En el núcleo celular se localizan unos corpúsculos, llamados cromosomas, cuyo
número, forma y tamaño varía según la especie. Los cromosomas son unos
filamentos, unos bastoncillos o hilos ubicados en el núcleo de las células
reproductoras y no reproductoras. Han recibido tal nombre por tener la
propiedad de teñirse con facilidad con cierto colorante, la cromatina. Los cromosomas
están constituidos por cadenas de moléculas de ADN. Cada gen ocupa un lugar
determinado dentro del cromosoma. Esa posición exacta se llama locus. Cada uno
de los dos genes del par de cromosomas se llama alelo. Si los dos alelos de un
mismo locus son idénticos, se dice que ese individuo es homocigótico. Si, en
cambio, los dos alelos son diferentes, el individuo es heterocigótico.
Las células de todos los seres vivos comprenden dos tipos de cromosomas: Un par
de cromosomas recibe el nombre de HETEROCROMOSOMAS o CROMOSOMAS SEXUALES y
todos los pares restantes se llaman AUTOSOMAS o CROMOSOMAS AUTOSÓMICOS. Estos
últimos son los no portadores de caracteres sexuales y encargados de transmitir
características morfológicas, fisiológicas, etc., de carácter hereditario.
En un cromosoma existen miles de genes, fragmentos de ADN responsables de los
caracteres del individuo, es decir, de sus rasgos fenotípicos.
En el canario existen 80 cromosomas dispuestos por parejas. Los cromosomas
están formados químicamente por ADN (ácido desoxirribonucleico) y proteínas.
Durante la división celular, los cromosomas aparecen divididos
longitudinalmente en dos cromátidas hermanas unidas por el centrómetro. De los
80 cromosomas, hay 18 que son de mayor tamaño (macrocromosomas), y los que
generalmente se tienen en cuenta para analizar la herencia genética. De estos
18 cromosomas, hay dos cromosomas sexuales (heterocromosomas), que determinan
el sexo del animal, y el resto (autosomas), que determinan las demás
características del animal.
El genoma es el conjunto de genes que posee un individuo, mientras que el
término genotipo hace referencia a los genes para uno o varios caracteres. El
conjunto de características que un ser vivo hereda de sus progenitores se
denomina GENOTIPO y aquellas que se aprecian exteriormente en él como
consecuencia de la transmisión del genotipo, se denomina FENOTIPO. Éste viene
determinado por el genotipo y las "condiciones ambientales"
(luminosidad, temperatura, alimentación, etc.) en las que se ha desarrollado.
Dicho de otra forma, la organización genética de un organismo, tanto respecto a
un locus como sobre todos sus genes colectivamente, es el genotipo. El genotipo
de un organismo le afecta, directa o indirectamente, en la manifestación de
rasgos a nivel molecular, físico y de comportamiento, en lo que se llama
fenotipo. En los locus de genes heterocigóticos (distintos), los dos alelos
interactúan entre sí para producir un fenotipo. Esta interacción puede a menudo
describirse como dominante o recesiva.
La herencia autosómica dominante es una de las distintas formas en que un rasgo
o trastorno se puede transmitir de padres a hijos. Si un rasgo o enfermedad es
autosómica dominante, quiere decir que la persona sólo necesita recibir el gen
anormal de uno de los padres para heredar el rasgo o la enfermedad. Existen dos
tipos de dominancia: Completa e incompleta. La dominancia completa permite que
los heterocigotos expresen el fenotipo mutante de manera absoluta. Con
dominancia incompleta los heterocigotos muestran un término medio entre el
fenotipo normal y el mutante.
La herencia autosómica recesiva es un rasgo o trastorno que deben estar
presentes dos copias del gen anormal, es decir, en el padre y en la madre para
que se desarrolle la enfermedad o el rasgo.
Si se cruza un canario melánico con uno lipocromo, se obtendrá un canario con
parte de su plumaje melánico y otra parte lipocromo, la presencia de melaninas
o lipocromo se presenta de forma separada en las diversas zonas del cuerpo, es
el llamado canario "pío". Esto sucede porque ambos genes tienen una
dominancia parcial respecto del otro, así ninguno de los dos se manifiesta de
forma completa. A este tipo de dominancia se le llama codominancia.
Si se cruza un canario amarillo con uno rojo, todas sus crías saldrán
anaranjadas, es decir, que la prole sale con una fusión de los rasgos de los
padres, sin presentar zonas de dominancia completa de alguno de los
progenitores. En este caso la herencia es una fusión de los genes de los padres
que se expande por todo el cuerpo sin presentar dominancia en ninguna de las
zonas del cuerpo. Este tipo de dominancia se llama dominancia intermedia o
semidominancia, y tiene en común con la codominancia que el aspecto físico
(fenotipo) de los hijos no es igual a ninguno de sus progenitores.

Antes de meterme con las Leyes de Mendel, os pongo algunos
conceptos básicos, como recordatorio:
• Gen: Es la unidad básica de
herencia o unidad microscópica de material hereditario ordenada linealmente,
que ocupa lugar definido en un cromosoma. Es el responsable de mantener las
características de la especie gracias a la información genética grabada en la
cinta del ADN. Cada ser tiene una cantidad de genes que son los que determinan
las características de éste. Los genes se agrupan en los organismos superiores
o cromosomas. Se pueden definir también como unidades, piezas o agentes
alojados en los cromosomas y que tienen por misión transmitir en el organismo
lo hereditario. Ellos son los responsables directos de la herencia con sus acciones.
• Diploide: Organismos que tienen
cromosomas iguales dos a dos, o lo que es lo mismo, contienen dos genes para
cada carácter.
• Genotipo: Es el contenido genético
del individuo en concreto, o dicho de forma más sencilla, todos los genes que
tiene.
• Fenotipo: Es la manifestación
visible de los genes, es decir, las cualidades físicas observables en un
organismo, incluyendo su morfología, fisiología y conducta a todos los niveles
de descripción.
• Cromosoma: Es el equivalente a un
paquete de genes. En cada cromosoma se agrupan una cantidad de genes. Cada
especie tiene un número determinado de cromosomas. La mitad de los cromosomas
son heredados de la madre y la otra mitad del padre.
• Locus: Ubicación del gen en un
cromosoma. Para un locus puede haber varios alelos posibles (plural: loci).
• Alelo: Es la pareja de genes,
procedente del padre y de la madre, que determinan un carácter. Es cada una de
las variantes de un locus, es decir, los dos genes de un mismo par de
cromosomas, procedentes del padre y de la madre. Cada alelo aporta diferentes
variaciones al carácter que afecta. En organismos diploides (2n) los alelos de
un mismo locus se ubican físicamente en los pares de cromosomas homólogos.
Son los genes que tienen la misma función, pero distintos efectos,
representando uno de ellos una modificación, una alteración del otro, aparecida
normalmente por causa de una mutación o algún debilitamiento. Uno se encuentra
alojado en un cromosoma y otro en el cromosoma homólogo, dándose el caso de que
ocupan el mismo lugar ("locus") y están al mismo nivel o
altura.
• Dominancia, Alelo dominante:
Predominio de la acción en un alelo sobre la de su alternativo (llamado alelo
recesivo), enmascarando u ocultando sus efectos. El carácter hereditario
dominante es el que se manifiesta en el fenotipo (conjunto de las propiedades
manifiestas en un individuo).
• Recesividad, Alelo recesivo:
Característica del alelo recesivo de un gen que no se manifiesta cuando está
presente el alelo dominante. Para que este alelo se observe en el fenotipo, el
organismo debe poseer dos copias del mismo alelo, es decir, debe ser homocigoto
para ese gen (según la terminología mendeliana, se expresaría como “aa”). En
caso contrario, se dice que el ejemplar es portador del carácter recesivo, pues
lo posee, aunque no lo manifiesta.
• Cariotipo: Composición fotográfica
de los pares de cromosomas de una célula, ordenados según un patrón estándar,
es decir, la fotografía de todos sus cromosomas. En un cariotipo se encuentra
el conjunto de características que permiten reconocer la dotación cromosómica
de una célula.
• Meiosis: Es el proceso de división
celular que permite a una célula diploide generar células haploides en
eucariotas. En este proceso se produce una replicación del ADN y dos
segregaciones cromosómicas, de manera que de una célula inicial diploide se
obtienen cuatro células haploides. Es el proceso que afecta a las células
germinales, que por división crean gametos con la mitad de cromosomas.
• Autosoma: Todo cromosoma que no
sea sexual.
• Gameto: Célula sexual que procede
de una estirpe celular llamada línea germinal. Tiene un número de cromosomas
haploides (n), debido al proceso de división celular (meiosis), que permite
reducir el número de cromosomas a la mitad. El gameto femenino se denomina
óvulo; el gameto masculino recibe el nombre de espermatozoide.
• Cigoto o huevo: Célula resultante
de la unión de dos gametos haploides. Es, por tanto, diploide (2n).
Generalmente, experimenta una serie de divisiones celulares hasta que se
constituye en un organismo completo. Su citoplasma y sus orgánulos son siempre
de origen materno al proceder del óvulo.
• Homocigoto: Individuo puro para
uno o más caracteres, es decir, que en ambos loci posee el mismo alelo
(representado como “aa” en el caso de ser recesivo o “AA” si es dominante). Se
dice que un gen está en homocigosis cuando está en el alelo procedente del
padre y en el alelo procedente de la madre en el mismo estado, es decir, mutado
o no mutado.

• Heterocigoto: Individuo que para
un gen, tiene un alelo distinto en cada cromosoma homólogo. Su representación
mendeliana es “Aa” o “aA”. Se dice que un gen está en heterocigosis cuando está
en el alelo procedente del padre y en el alelo procedente de la madre en
estados diferentes, es decir, uno de ellos está mutado y el otro no mutado.
• Híbrido: Es el resultado del
cruzamiento o apareamiento de dos individuos puros homocigotos (uno de ellos
recesivo y el otro dominante) para uno o varios caracteres.
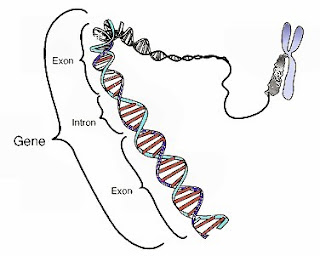
En el ADN del cromosoma existen fragmentos que llevan información para la
síntesis de proteínas y son llamados exones. Entre estos fragmentos se
intercalan otros fragmentos llamados intrones, que no almacenan información
para la síntesis de proteínas y, por otra parte, incrementan la distancia entre
los exones, lo que aumenta la posibilidad de crossing-over y con ella, la
variabilidad genética en los canarios.
En genética, cuando se representan los genes con letra
mayúscula, significa que son genes dominantes. Si se representan con letra
minúscula, significa que son genes recesivos.
• Primera Ley; Ley de la Uniformidad de los Híbridos de la primera generación:
Si se cruzan dos individuos de raza pura (homocigóticos) y que difieren en uno
o más caracteres, toda la descendencia de primera generación (F1) será igual,
es decir, el mismo tipo de híbrido, con igual fenotipo e igual genotipo, e
iguales (en fenotipo) a uno de sus progenitores. Como ilustra el dibujo se
puede observar que los guisantes amarillos de raza pura tienen los dos genes
"A", mientras que los verdes los tienen "a". Los
descendientes pese a que tienen tanto el gen "A" como el
"a" sólo expresan el "A", puesto que es el dominante, es
decir, la descendencia es amarilla.

Tercera Ley; Ley de Herencia Independiente o Ley de la
Independencia de Caracteres: Dos caracteres diferentes se heredan y se
comportan de forma independiente para cumplir cada uno con la primera y segunda
ley, es decir que no influye un carácter sobre otro a la hora de transmitirse. Mendel
observó esto en los guisantes cuando, además de en el color, se fijó en que
existían guisantes lisos y rugosos. El gen dominante es el de guisante liso
"B" mientras que el del rugoso "b" es recesivo. Al
cruzarlos comprobó que los caracteres se transmiten de forma independiente.

Además, hay una serie de excepciones a la 3ª Ley de Mendel:
La herencia monogenética o mendeliana es el tipo de transmisión que más afecta
a las aves, sin que se excluya la posibilidad de que existan interacciones
entre genes, cada uno responsable por sí mismo de la aparición de un efecto
fenotípico concreto.
La transmisión independiente de los caracteres no siempre se cumple, es decir,
que muchos de ellos se transmiten juntos en la descendencia, al estar situados
los genes para distintos caracteres en el mismo cromosoma.
Cuando los genes están ligados, la 3ª ley de Mendel no se cumple, a no ser que
se produzca entrecruzamiento (crossing-over) o intercambio genético. Por
ejemplo, el gen pastel está situado sobre el cromosoma sexual Z, al igual que
el gen para el color marfil. El gen no pastel y el gen no marfil son alelos
respectivamente de los genes pastel y marfil. Como se encuentran situados en un
mismo cromosoma genes para distintos caracteres (pastel- marfil, no pastel-no
marfil), se dice que están ligados y, en este caso, ligados al sexo.
Ya que durante la meiosis se separan los cromosomas homólogos, es evidente que
el gen marfil permanece unido al gen pastel, y lo mismo ocurrirá con los genes
no pastel y no marfil. Es decir, el color pastel se acompañará siempre de la
mutación marfil.
En definitiva, todos los genes situados en un mismo cromosoma se transmiten
ligados o asociados, no pudiendo separarse ni recombinarse independientemente.
La 3ª ley de Mendel en tales casos no se cumplirá, salvo que ocurra
entrecruzamiento. En el caso, por ejemplo, de los genes para la categoría
intenso y nevado, y los genes para el carácter marfil sí se cumplirá, ya que se
localizan estos genes en cromosomas diferentes, uno autosómico y el otro
sexual.
La determinación del sexo está regulada por esos cromosomas
especiales, llamados cromosomas sexuales. También reciben el nombre de
heterocromosomas porque son diferentes morfológicamente y poseen un contenido
génico distinto.
En las aves en general, el par de cromosomas sexuales de la hembra se compone
de un cromosoma de tamaño normal (Z) y de otro de tamaño reducido (W),
atrofiado, con una morfología (forma) distinta a la de su normal homólogo. A
este cromosoma W solo se le reconoce hasta la fecha el alojamiento del gen que
trasmite el carácter sexo hembra, que es un carácter más, pues, es algo que se
hereda y que produce los rasgos propios de la hembra. La cría, que recibe de la
madre tal cromosoma atrofiado, es hembra. El cromosoma normal Z es idéntico a
los que componen el par de cromosomas sexuales del macho al ser del mismo
tamaño, la misma forma y poder alojar los mismos genes. Los dos del macho y el
normal de la hembra se consideran activos por alojar genes distintos al
carácter del sexo. A los dos cromosomas sexuales del macho se les llama
"ZZ" y los de la hembra se los nombra por "ZW".
Hay algunos caracteres que están determinados por genes que se encuentran en
los cromosomas sexuales y, por tanto, se heredan a la vez que el sexo. El tipo
de herencia de estos caracteres se denomina herencia "ligada al
sexo". Cuando una mutación surge en el cromosoma Z, su transmisión es
“ligada al sexo”. Todas las mutaciones hoy conocidas del cromosoma Z son
recesivas en relación a su original. Los genes ligados al sexo tienen un
comportamiento distinto en los machos y en las hembras. Ello es debido a que el
cromosoma Z es diferente del cromosoma W.
Se dice que el sexo homogamético en las aves es el masculino, ya que contiene
dos cromosomas iguales ZZ, y el sexo heterogamético es el femenino, ya que
contiene un cromosoma sexual Z y otro W. Los espermatozoides portan siempre el
cromosoma Z, mientras que los óvulos llevan el cromosoma Z en un 50% y el W en
otro 50%. De esta forma, al formarse los cigotos tras la fecundación, el 50%
será hembras (ZW) y el otro 50% machos (ZZ). El coeficiente sexual, es decir,
la proporción de sexos es igual a 1, siendo el cromosoma Z el transmisor del
factor ligado al sexo.

En los machos, que poseen dos cromosomas ZZ, puede haber heterocigosis y, en
ese caso, el fenotipo expresará la característica del alelo dominante. Como
todas las mutaciones ligadas al sexo son recesivas en relación al original, en
los machos el efecto de ellas sólo se hará sentir en el fenotipo cuando se
presente en homocigosis.
Hay que destacar que es suficiente con que un macho sea portador de un gen
recesivo ligado al sexo, para obtener hijas hembras con este factor. Al tener
las hembras un solo cromosoma Z, nunca pueden ser portadoras de un factor
recesivo y ligado al sexo, ya que los genes recesivos se expresan, aunque estén
en un único gen, dada la falta del alelo en el cromosoma sexual homólogo. Como
el cromosoma W de las hembras no posee genes, todos los genes presentes en su
cromosoma Z expresan sus efectos en el fenotipo. Por tanto, las hembras nunca
pueden ser portadoras de factores ligados al sexo; o son mutadas o son
clásicas.
En el caso de los genes del cromosoma Z, la transmisión presenta una
particularidad en relación a los autosómicos: Depende del sexo de los
progenitores.
Con frecuencia, en la cría aparecen algunos ejemplares con características que
en su fenotipo no mostraban los progenitores. Esto puede ser debido a dos
posibilidades:
1.- Que los dos progenitores eran portadores del carácter.
2.- Que el carácter en cuestión es ligado al sexo; en este caso, se puede
afirmar:
a).- El progenitor macho era portador del carácter.
b).- Que dichos ejemplares son hembras.
Las mutaciones ligadas al sexo son el negro-bruno clásico, bruno, ágata,
isabela, satiné, lutino, pastel, marfil y eumo (sólo en el jilguero, ya que en
el canario la mutación eumo es autosómica recesiva).
Los caracteres influidos por el sexo son aquellos que se manifiestan de forma
diferente según el sexo del individuo, ello es debido a la acción de hormonas
sexuales que influyen sobre la manifestación de ese carácter. El gen mosaico es
un ejemplo, así como la presencia cuantitativa de feomelanina, que es mayor en
la hembra. También la cantidad de pigmento lipocrómico, el factor óptico de
refracción, la expresión de las categorías, intenso y nevado, son caracteres
determinados por genes cuya expresión fenotípica depende del sexo del
individuo.
La Consanguinidad
La Consanguinidad es la relación de parentesco que tiene
lugar entre dos individuos. Se habla de cruzamiento consanguíneo o endogamia
cuando se cruzan entre sí dos individuos con relaciones de consanguinidad.
La consanguinidad, especialmente si es repetitiva, provoca taras, disminución
del tamaño y degeneración genética. Es útil para fijar caracteres recesivos,
por ejemplo, surgidos tras una nueva mutación, pero también contribuirá a fijar
los defectos que, a partir de entonces, siempre aparecerán en sus
descendientes. Por la consanguinidad tienen más posibilidades de aparecer en la
descendencia genes letales recesivos, al combinarse en los progenitores
emparentados. Reproductores que poseen una dotación genética común pueden más
fácilmente determinar el nacimiento de una prole enferma, como consecuencia del
hecho de que aumenta la posibilidad de que ambos reproductores sean portadores
de taras hereditarias latentes, es decir, que genes con afección de carácter
recesivo se manifiesten precisamente gracias a la unión de dos portadores.
La máxima consanguinidad tiene lugar en el cruce de hermanos (100%), padres-
hijos (50%). Teniendo presente que debe evitarse el cruce entre hermanos, la
unión entre consanguíneos en perfectas condiciones físicas no produce en
general grandes inconvenientes, siempre que se tenga la precaución de no
insistir en él, introduciendo con frecuencia sangre nueva en el criadero.
Los acoplamientos consanguíneos en línea estrecha deben basarse en cruzamientos
en los que prevalezca en forma absoluta la sangre de uno de los reproductores,
es decir, no pueden ser fruto de una exasperada e irracional insistencia en una
sola dirección, debiendo procederse con método en base a los resultados
obtenidos por la experiencia, con la idea de buscar un fin óptimo o de fijar un
carácter nuevo. Si el resultado se desvía del objetivo buscado, hay que
abandonar y no insistir en la mala dirección.
A continuación os pongo y explico el esquema de cría en consanguinidad de
Felch, para un proceso de cría que lleva de 5 a 6 años:

En el primer año de cría, de macho (1) y hembra (2) seleccionados como origen
de progenie consanguínea se obtienen sujetos "media sangre" (3). Esta
prole (3) o "media sangre" son los sujetos en los cuales la
conformación genética está influenciada en igual medida por la sangre paterna
como la materna (50/50). A estos sujetos es necesario recurrir durante las
sucesivas uniones consanguíneas para quebrar la absoluta primacía de una de las
dos líneas ascendentes.
El segundo año de cría, el criador debe elegir las mejores hembras para unirlas
al padre y el mejor macho para acoplar con la madre. Seleccionar dos hembras
del grupo (3) y aparearlas con el macho original (1), produciendo así el grupo
(4). Además, el mejor macho del grupo (3) cruzarlo con la hembra original (2),
obteniendo así el grupo (5). Estos dos grupos colaterales (4) y (5) tendrán una
dominancia de la sangre paterna o materna en el porcentaje 75/25.
El tercer año de cría, considerando que el estado de salud y el vigor de los
ejemplares lo permitan, la mejor hembra del grupo (4) aparearla con el macho
original (1); ésto producirá el grupo (6). El mejor macho del grupo (5)
aparearlo con la hembra original (2); ésto nos dará el grupo (8). El resultado
serán sujetos con 7/8 de sangre paterna y 1/8 de sangre materna (6) y otros con
inversa proporción sanguínea (8).
Aquí surge nuevamente la oportunidad de obtener "media sangre",
seleccionando también el mejor macho del grupo (4) y aparearlo con una buena
hembra del grupo (5), obteniendo así el grupo (7). El mismo cruce se puede
hacer en sentido opuesto, es decir, un buen macho del grupo (5) con una buena
hembra del grupo (4), produciendo también el grupo (7).
Los sujetos de los grupos (6) y (8) pueden ser emparejados con los
ascendientes directos (4 y 5) y también con los "media sangre".
El cuarto año se pueden hacer los siguientes cruces: El grupo (4) con el grupo
(6) para obtener el grupo (9). El grupo (5) con el grupo (7) para obtener el
grupo (10). El grupo (6) con el grupo (8) para obtener el grupo (11). El
grupo (7) con el grupo (8) para obtener el grupo (12). Y el grupo (8) con
el grupo (5) para obtener el grupo (13).
El resultado serán ejemplares (grupos 9 y 13) poseedores de 13/16 sangre de uno
de los progenitores y 3/16 sangre del otro, y ejemplares (grupos 10 y 12) cuya
desproporción entre las calidades de sangre resultará atenuada, con 11/16 y
5/16 con preeminencia de uno y otro. Además, se sigue disponiendo de “medias
sangre” en el grupo (11).
El quinto año se pueden hacer los siguientes cruces: El grupo (9) con el grupo
(11) para obtener el grupo (14). El grupo (9) con el grupo (12) para obtener el
grupo (15). El grupo (10) con el grupo (12) para obtener el grupo (16). El
grupo (9) con el grupo (13) para obtener también el grupo (16). El grupo (10)
con el grupo (13) para obtener el grupo 17. Y, finalmente, el grupo (11) con el
grupo (13) para obtener el grupo (18).
Llegados a este punto, conviene truncar la operación "proliferación
consanguínea" e introducir sangre fresca, recurriendo a un sujeto
apropiado de otra cepa, genéticamente puro, que tenga características
homocigóticas que se transmiten fielmente a la descendencia.
Si en lugar de este ejemplar puro, se introduce un reproductor heterocigoto,
podrían anularse muchos años de esfuerzos selectivos a causa de la transmisión
de características que el sujeto no presenta y de los que es portador.
La estrecha reproducción consanguínea que se ha ilustrado en la Tabla de Felch
permite mantener íntegras las buenas características a la descendencia.
Observando el cuadro, se puede deducir que cuando se han obtenido los grupos 3,
4, 5, 6, 7 y 8 con éxito, el criador cuenta ya con un número suficiente de
pájaros de cada grupo y, por consiguiente, los servicios de los dos pájaros de
origen ya no son necesarios para el futuro proceso del sistema. Se desprende de
aquí que debe tenerse especial cuidado con la pareja de origen, ya que su uso
es indispensable hasta el tercer año.
EPISTASIA.
Cuando un gen enmascara y anula a otro gen no alélico a él, se dice que se ha
producido una epistasia. La epistasia o interacción génica se presenta cuando
un solo carácter depende de dos o más pares de genes, de forma que la
manifestación de un par de genes depende de la información de otro par de genes
u otros pares de genes. Por esta interacción, los alelos que enmascaran a otros
alelos reciben el nombre de epistáticos y los que quedan anulados o
enmascarados, hipostáticos. De la misma forma, al gen que suprime la
manifestación de otro se le llama gen epistático, y al suprimido, gen
hipostático. Los genes epistáticos pueden ser dominantes o recesivos.
La epistasia se observa siempre en los cruces heterocigóticos, es decir, en los
cruces de los F1, y se manifiestan sus resultados en los descendientes o F2. El
gen epistático puede ir perfectamente combinado con el gen dominante y recesivo
mendeliano.
Un ejemplo de epistasia en el canario es el gen blanco recesivo, que en
homocigosis es epistático con relación a los genes para el color amarillo
marfil, rojo marfil, blanco dominante, amarillo y rojo. Es decir, un canario
blanco recesivo puede ser portador de amarillo, rojo, marfil, blanco dominante,
etc., pues el gen para el blanco recesivo no es alelo de ninguno de los genes
citados.
La epistasia hace variar las proporciones clásicas de la 3ª ley de Mendel
(9:3:3:1), de forma distinta según el tipo de epistasia. En el caso de que el
gen epistático fuese recesivo, como es el gen blanco recesivo, las proporciones
quedarían modificadas en 9:3:4.
Por otro lado, se conoce que el gen blanco dominante es epistático incompleto
(no inhibe completamente la manifestación de otras parejas de genes) con
relación al gen marfil. En este caso, la epistasia es de tipo simple dominante
y se modifica la proporción mendeliana a 12:3:1, pues un canario genéticamente
marfil, no expresa dicho genotipo mas que de forma parcial, cuando esté
además presente el gen blanco dominante.
En resumen, el gen blanco recesivo es epistático con relación al gen blanco
dominante, que a su vez es epistático con relación al gen marfil. Se podría
hablar en este caso de reepistasia y de genes reepistáticos. La reepistasia
supone la existencia de tres genes distintos, cada uno con sus correspondientes
alelos, localizados en tres loci diferentes, pertenecientes o no al mismo
cromosoma, y entre los cuales se da un orden de jerarquía. La epistasia
del gen blanco dominante sobre los genes amarillo y rojo, parece ser que
también tiene lugar.
Las clases de epistasia con interacción de dos factores y los resultados
genotípicos y fenotípicos de los F2, son los siguientes:
Intragénica clásica mendeliana..……….9:3:3:1
Epistasia simple recesiva……………….9:3:4 (única en canaricultura).
Epistasia simple dominante…………….12:3:1
Epistasia con efectos acumulativos……9:6:1
Epistasia doble recesiva……………….. 9:7
Epistasia doble dominante……………...15:1
Epistasia doble dominante-recesiva…...13:3
Para simplificar la búsqueda de una interacción genética hay que tener en
cuenta, la cantidad de fenotipos que se manifiestan: Si son CUATRO, se trata de
la herencia mendeliana; si fueran TRES fenotipos, serían epistasias simples
recesivas o simples dominantes o con efectos acumulativos; y si los fenotipos
que se produjeran fueran DOS, se trataría de epistasias dobles recesivas,
dobles dominantes o dobles dominante-recesiva.
En el caso particular de la epistasia simple recesiva (F2.- 9:3:4), única en
canaricultura, se produce cuando en un gen su alelo recesivo homocigótico “aa”
es epistático. En este caso, anula a otro gen o genes no alélicos dominantes.
El caso más característico es el del pájaro heterocigótico blanco semidominante
“AB”, para el amarillo y el blanco, cruzado con el amarillo portador del blanco
recesivo “Aa”. Los gametos masculinos y femeninos que originan tras la
fecundación estos F1 son: “AA”, “AB”, “aA” y “aB”.

Como se puede observar en la tabla, el resultado de esta combinación es de 9
genotipos y fenotipos blancos, (6 semidominantes portadores de blanco recesivo
y 3 blancos semidominante); 3 amarillos (1 homocigótico y dos portadores) y 4
blancos recesivos, por anular el alelo recesivo epistático “aa” a los alelos
“AA”, “AB”, “BA” y “BB”, que son los hipostáticos.
Con este mismo resultado se podrían hacer las fórmulas para el cruce de los
distintos lipocromos, y aplicado a los melánicos, aquellos que tengan de color
de fondo el blanco recesivo (gris recesivo, cobalto, onix, bruno plata, pastel
plata, satiné plata, etc.). Todos ellos llevarán en su genotipo los genes para
los lipocromos amarillo, rojo, o blanco semidominante.
Con el cruce de dos F2 blancos recesivos para obtener F3 se demuestra que los
genes recesivos homocigóticos son epistáticos y anulan a los genes amarillos,
rojos y blanco semidominante, o sea que la casilla que tenga los alelos “aa”
serán blancos recesivos anulando a los genes acompañantes, permitiendo
solamente la presencia del blanco recesivo, ya sea como lipocromo o como fondo
de los melánicos.
Se escogerá para la prueba el cruce “aaBA” por “aaBA” de dos blancos recesivos,
que producen los siguientes gametos: “aB”, “aA”, “aB” y “aA”, para el macho y
para la hembra.

HIBRIDACIÓN.
Aunque la hibridación del canario con otros fringílidos no es exactamente un
carácter influido por el sexo, sí se ve afectado por el mismo ya que, según el
tipo de hibridación, toda la descendencia será estéril o sólo las hembras. Al
igual que en los canarios heterocigóticos, al híbrido producto del cruzamiento
de ejemplares de dos especies distintas se le denomina F1. Así los
descendientes de jilguero x canaria, son los F1 del cruce. Este cruce acaba en
los F1 ya que los descendientes son estériles. Pero si se cruzan dos especies
diferentes y los descendientes son fértiles, se podrá disponer de F1 fértiles
y, por tanto, cruzándolos entre sí se llegará a individuos F2 que, cruzándolos
entre sí darán F3, y así sucesivamente.
Canario x verdecillo = F1 de canario por verdecillo
F1 x F1 = F 2 de canario por verdecillo
F2 x F2 = F 3 de canario por verdecillo
F3 x F3 = F4 de canario por verdecillo…
Para evitar la consanguinidad, los F1 pueden venir de parejas diferentes, pero
que tienen las mismas características.
Cuando se dispone de un pájaro híbrido que sea fértil, si lo cruzamos con un
ejemplar de la misma raza que su ancestro canario o su ancestro silvestre, se
le denomina cruzamiento retrógrado y a la descendencia se les denomina R1.
Esta marcha atrás se puede hacer con uno de los dos padres indicando si es para
el canario o para el silvestre. Como todas las hembras suelen ser estériles, se
debe emplear el F1 fértil como macho y la hembra del ancestral hacia el que se
quiera tender.
De este primer cruce los R1 tendrán un 75% del ancestral hacia el que se quiere
tender y un 25% del otro. Las hembras R1 serán de escasa fertilidad y los
machos no con la fertilidad completa. Al cruzar el macho R1 con otra hembra del
ancestral hacia el que se quiere tender, se obtendrán R2 con un 87,50% del
ancestral buscado y un 12,50% del otro. Esta descendencia R2 será de machos
fértiles y hembras con fallos en la reproducción. Al cruzar el macho R2 con
otra hembra del ancestral hacia el que se quiere tender, se obtendrán R3 con un
93,75% del ancestral buscado y un 6,25% del otro. Esta descendencia R3 será de
machos y hembras fértiles.
F1 de canario por verdecillo x verdecillo = R 1 hacia verdecillo
F1 de canario por verdecillo x canario = R 1 hacia canario.
R1 x verdecillo = R2 hacia verdecillo
R2 x verdecillo = R3 hacia verdecillo...
MUTACIONES:
Cuando la herencia no se transmite por el sistema habitual de duplicación,
puede producirse súbitamente un error o “accidente" en la cinta del ADN.
En este caso, aparece un ser vivo con cualidades fenotípicas o genotípicas
distintas de las de sus progenitores.
La mutación espontánea o natural es una alteración en la secuencia del ADN,
producida por errores en la replicación del ADN, reparación incorrecta de dicho
ADN o recombinación desigual, que desembocan en un cambio en la información
genética (genotipo) de un ser vivo y que, por lo tanto, va a producir un cambio
de características, que se presenta súbita y espontáneamente, y que se puede
transmitir o heredar a la descendencia. La unidad genética capaz de mutar es el
gen que es la unidad de información hereditaria que forma parte del ADN. En los
canarios, como seres multicelulares que son, las mutaciones sólo pueden ser
heredadas cuando afectan a las células reproductivas. Pueden ocurrir en las
células somáticas, afectando sólo al individuo en el que ocurren, o bien,
afectan a las células germinales, siendo transmitidas a la descendencia.
Si se examina el color grisáceo de un canario silvestre, se puede encontrar una
mezcla de pigmentos marrones y negros sobre un color base amarillo. Si por
algún motivo (accidente genético) se altera la información existente en un gen,
éste no podrá suministrar la información que poseía y suministrará otra
diferente. Estos accidentes genéticos son mutaciones, aberraciones cromosómicas
o alteraciones genéticas (cambio genético repentino en un gen o grupo de
genes). Son cambios inesperados en la información genética de un ser vivo, no
debidos a la segregación ni recombinación genéticas. Generalmente, las
mutaciones son de carácter recesivo y desfavorable.
La causa de este proceso, en la mayoría de los casos, no se conoce, pero se
sabe que algunas mutaciones pueden aparecer en las células sexuales de un
animal que está siendo sometido a algún tipo de radioactividad. Las mutaciones
pueden aparecer espontáneamente (mutaciones espontáneas o naturales) o ser
inducidas por agentes mutágenos (mutaciones experimentales, inducidas o
artificiales). Entre los agentes mutágenos se encuentran los rayos X, rayos UV,
radiaciones atómicas, ultrasonidos, bromouracilo, etc.
La mayoría de las mutaciones no resultan prácticas para el criador de canarios
y, en muchos casos, ni siquiera son percibidas por él. No alteran
necesariamente un color ni la forma exterior. Las mutaciones importantes para
el criador son aquellas que afectan al color de las plumas o, en un menor
grado, a la forma general del canario y de su plumaje.
Es fácilmente comprensible lo importante que han sido las mutaciones para
desarrollar todas las variedades diferentes de canarios. Los criadores también
emplean las mutaciones para su propio beneficio, es decir, para mejorar las
distintas razas. Las mutaciones son esenciales a la hora de desarrollar colores
y formas completamente diferentes. Al cruzar una mutación con otra, se pueden
producir más variedades. Teóricamente no hay límite con relación al número de
variedades que se pueden obtener, siempre que las mutaciones continúen
apareciendo regularmente.
En los cromosomas existen algunos genes que, al unirlos, se consideran letales,
ya que provocan el aborto de embriones o la muerte de los individuos que los
presentan. En el canario de color se conocen dos genes dominantes (blanco
dominante e intenso), que se comportan como letales en homocigosis.
Por la pérdida de los pigmentos negro y marrón apareció el canario amarillo. El
mismo canario amarillo tiene muchas más posibilidades. En primer lugar porque
es mucho más atractivo que sus parientes grises, pero también porque tiene
muchas posibilidades genéticas.
En resumen, las mutaciones implican nuevas variedades, la selección abre el
camino a mejores especímenes, el cruce de mutaciones produce más variedades de
color y se pueden producir todavía más colores por medio de la hibridación (de
esta forma el canario de factor rojo logró su color gracias al Cardenalito de
Venezuela).
La tasa de mutación (Tm) es la frecuencia con la que un alelo silvestre
presente en un locus cambia a un alelo mutante. Se expresa como el número de
mutaciones por unidad biológica (división celular, gameto o ronda de
replicación). Los factores que afectan a la tasa de mutación son tres:
Ø La frecuencia del cambio
en el ADN (espontáneo o inducido).
Ø La probabilidad de que un
cambio en el ADN sea reparado.
Ø La probabilidad de
detectar la mutación.
La tasa de mutación es escasa (una por cada millón de gametos) y depende del
gen de que se trate. Hay genes que mutan con facilidad (genes inestables)
mientras que otros lo hacen con una frecuencia muy escasa (genes estables). Una
vez producida la mutación, el gen mutante es tan estable e invariable como el
del que procede (gen silvestre).
En el canario se puede comprobar que, en un tiempo relativamente corto de cría
en cautividad, se han producido, detectado y fijado muchas mutaciones. A pesar
de todo, hay que tener en cuenta que un gran porcentaje de esas mutaciones se
han perdido o han pasado desapercibidas. La mayor parte de las mutaciones que
se han conseguido fijar afectan a un solo par de cromosomas (heterocromosomas o
cromosomas sexuales) y que, por este motivo, permite una mayor facilidad de
detección al mostrarse en el fenotipo de las hembras que posean la
característica. Si se considera que un número similar de alteraciones han
ocurrido en el resto de los 39 pares de macrocromosomas (autosomas o cromosomas
autosómicos), una elemental operación matemática dará idea de las que se han
perdido o han pasado desapercibidas.
Las mutaciones pueden ser inocuas, beneficiosas o desfavorables. Generalmente
son recesivas y desfavorables, pudiendo producir determinadas enfermedades y
taras genéticas, e incluso en algunos casos, la muerte. Por lo general, los
ejemplares mutados suelen ser más débiles, y no es muy conveniente la
reiteración indefinida en su apareamiento. Si la mutación afecta a un carácter
dominante, se localiza con facilidad; en cambio, si es recesivo resulta más
difícil su detección, ya que sólo se manifiesta en homocigosis.
Las mutaciones pueden provocar aumento o disminución del número de cromosomas
(aneuploidía), juego de cromosomas (euploidía), pérdida de información
genética, etc. Según un estudio reciente, los tipos de mutaciones más
frecuentes en aves son las inversiones, translocaciones y triploidías.
Las inversiones se deben a giros de 180º en la posición de algunos genes en el
cromosoma, siendo el resultado un cromosoma que contiene un segmento en orden
inverso al normal.
Las translocaciones ocurren cuando dos segmentos del mismo cromosoma
(intracromosómicas) o de dos cromosomas (intercromosómicas) cambian su lugar el
uno con respecto al otro y viceversa.
La triploidía consiste en que se presentan tres juegos cromosómicos (3n) en
lugar de dos (2n, diploidía), dado que, por error, un individuo recibe de un
progenitor dos copias de cada cromosoma, en lugar de una.
Todas estas mutaciones provocan disminución en la fertilidad.
Por último, un apunte sobre los factores de carácter recesivo. Son aquellos
que, para obtener ejemplares con este factor, es necesario como mínimo que
ambos progenitores sean portadores del factor o mutación. Por tanto, para que
el factor se exprese externamente debe haber homocigosis, es decir, todos los
genes que afectan a este carácter deben ser iguales en el individuo afectado.
Estos factores de carácter recesivo se localizan en los cromosomas autosómicos,
por consiguiente, tanto el macho como la hembra pueden ser portadores. Los
principales factores autosómicos de carácter recesivo son el blanco recesivo,
opal, phaeo, topacio, eumo y onix.
ADN...
El ADN tiene la función de “guardar información”. Es decir, contiene las
instrucciones que determinan la forma y características de un organismo y sus
funciones. Además, a través del ADN se transmiten esas características a los
descendientes durante la reproducción, tanto sexual como asexual. Todas las
células, procariotas y eucariotas, contienen ADN en sus células. En las células
eucariotas el ADN está contenido dentro del núcleo celular, mientras que en las
células procariotas, que no tienen un núcleo definido, el material genético
está disperso en el citoplasma celular.

El ADN está organizado en cromosomas. En las células eucariotas los cromosomas
son lineales, mientras que los organismos procariotas, como las bacterias,
presentan cromosomas circulares. Para cada especie, el número de cromosomas es
fijo. Por ejemplo, los canarios tienen 80 cromosomas en cada célula, agrupados
en 40 pares, de los cuales 39 son autosomas y un par es sexual (ZZ o ZW).
Cada cromosoma tiene dos brazos, ubicados por arriba y por debajo del
centrómero. Cuando los cromosomas se duplican, previo a la división celular,
cada cromosoma está formado por dos moléculas de ADN unidas por el centrómero,
conocidas como cromátidas hermanas.
El ADN se compone de dos cadenas, cada una formada por nucleótidos. Cada
nucleótido, a su vez, está compuesto por un azúcar (desoxirribosa), un grupo
fosfato y una base nitrogenada. Las bases nitrogenadas son cuatro: adenina (A),
timina (T), citosina (C), y guanina (G), y siempre una A se enfrenta a una T y
una C se enfrenta a una G en la doble cadena. Las bases enfrentadas se dice que
son complementarias.

El ADN adopta una forma de doble hélice, como una escalera caracol donde los
lados son cadenas de azúcares y fosfatos conectadas por “escalones”, que son
las bases nitrogenadas. La molécula de ADN se asocia a proteínas, llamadas
histonas, y se encuentra muy enrollada y compactada para formar el cromosoma.
Esta asociación de ADN y proteínas se conoce como cromatina. La cromatina puede
estar enrollada en mayor o menor grado, dependiendo de la etapa en que se
encuentra la célula; por ejemplo, cuando el ADN se ha duplicado antes de que la
célula se divida, la cromatina se compacta en su mayor grado, y como resultado
se pueden visualizar los cromosomas duplicados al microscopio como corpúsculos
con forma de X.
Cuando la célula se divide, cada nueva célula que se forma debe portar toda la
información genética, que determine sus características y funciones. Para
eso, antes de dividirse, el ADN debe replicarse, es decir, generar una copia de
sí mismo. Durante la replicación, la molécula de ADN se desenrolla, separando
sus cadenas. Cada una de éstas servirá como molde para la síntesis de nuevas
hebras de ADN. Para eso, la enzima ADN-polimerasa coloca nucleótidos siguiendo
la regla de apareamiento A-T y C-G. El proceso de replicación del ADN es
semiconservativo, ya que al finalizar la duplicación, cada nueva molécula de
ADN estará conformada por una hebra “vieja” (original) y una nueva.

La información está guardada en el ADN en el código de secuencia de bases A, T,
C y G que se combinan para originar “palabras” denominadas genes. Los genes son
fragmentos de ADN cuya secuencia nucleotídica codifica para una proteína. Es
decir, que a partir de la información “escrita” en ese fragmento de ADN se
fabrica (sintetiza) un tipo particular de proteína. Aunque, en realidad, los
genes también llevan la información necesaria para fabricar moléculas de ARN
(ribosomal y de transferencia) que intervienen en el proceso de síntesis de
proteínas. El ARN (ácido ribonucleico) es una molécula con una estructura
similar al ADN pero, en vez de ser una doble cadena, es una cadena sencilla,
que transporta la información contenida en el ADN.
A veces, y este es un fenómeno relativamente frecuente, la enzima que se
encarga de la replicación del ADN (ADN polimerasa) se equivoca, es decir,
coloca un nucleótido en lugar de otro. Si, por ejemplo, la enzima ADN
polimerasa coloca una T en lugar de una A podría ocurrir que al traducirse, se
coloque en la proteína un aminoácido diferente del que correspondería. Por lo
tanto, la proteína generada sería diferente en un aminoácido a la original.
Este cambio en el ADN, llamado mutación, podría alterar o anular la función de
la proteína.
Este ejemplo ilustra el efecto de los cambios o mutaciones puntuales (debidos a
un único cambio en la secuencia) en la proteína final. En algunos casos las
mutaciones pasan inadvertidas, pero también pueden provocar la falta de
actividad de una proteína esencial y causar una enfermedad. De todas formas, la
mayoría de las mutaciones no se manifiestan, o porque están en regiones del ADN
donde no hay genes, o porque no cambian el aminoácido, o porque ese cambio no
altera la función de la proteína. O bien podría alterarse la función y esto no
resultar perjudicial. Tal es el caso del carácter color de ojos, donde el color
claro se produce por falta de ciertas enzimas que fabrican los pigmentos del
iris.
En realidad, las mutaciones son la base de la biodiversidad. Es decir,
que las pequeñas diferencias en el ADN es lo que determina que los seres vivos
sean diferentes entre sí. Esta diversidad en las características, sumada a la
existencia de un código genético común entre los seres vivos, son dos hechos
determinantes en el desarrollo de la biotecnología moderna.
"CROSSING OVER",
responsable de muchas mutaciones genéticas...
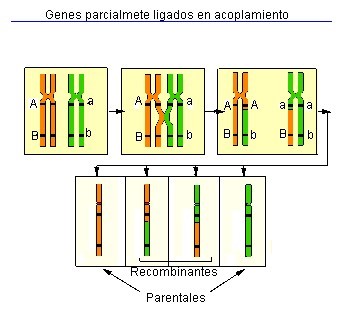
Hay una tendencia a que todos los genes de un mismo cromosoma, cualquiera que sea,
se transmitan juntos. Este fenómeno es conocido por ligación factorial o
“linkage”. Este fenómeno es más fuerte cuanto más próximos están los genes unos
de otros. Sin embargo, al unir cromosomas, puede producirse un intercambio de
fragmentos homólogos de un cromosoma al otro (intercambio de secciones de su
ADN). A esto se le denomina “crossing over” o entrecruzamiento cromosómico y
permite nuevas combinaciones de genes, ya que se produce intercambio de
material genético en el momento de la "meiosis".
Antes de iniciarse la meiosis, durante la interfase, ha ocurrido la duplicación
del ADN y cada cromátida da lugar a una cromátida homóloga, que quedan unidas
por el centrómero. Comienzan entonces las dos divisiones sucesivas. La primera
división meiótica consta de cuatro fases: Profase I, metafase I, anafase I y
telofase I. Es durante la profase I de la primera división meiótica cuando
tiene lugar el fenómeno del sobrecruzamiento cromosómico o crossing-over. Esta
fase es la etapa más larga y en la que se dan los acontecimientos más
característicos de la meiosis. En esta fase se constituyen los cromosomas al
desespiralizarse el ADN. Los cromosomas homólogos se juntan y entre ellos tiene
lugar un intercambio de fragmentos de ADN, es decir, los cromosomas homólogos
se emparejan y tiene lugar el intercambio del material hereditario. Los pares
de cromosomas homólogos están estrechamente apareados y se adhieren en
determinados puntos, denominados quiasmas. En esta situación, las cromátidas
hermanas se entrecruzan y se fragmentan transversalmente, dando lugar a un
intercambio de ADN entre ellas. La consecuencia de este intercambio es la
recombinación genética.

Explicándolo de una forma más simple, el crossing-over es un mecanismo por el
que dos cromosomas homólogos pueden intercambiar una secuencia de genes del
mismo tamaño que contenga los mismos “loci” y se recombinen del mismo modo que
su combinación de genes (alelos). Esto ocurre durante la meiosis, fase donde
las cromátidas hermanas se forman. Esto puede ocurrir varias veces, produciendo
entrecruzamiento múltiple. El lugar de unión donde se produce el crossing-over
es el quiasma y en un mismo cromosoma puede haber varios quiasmas. La
frecuencia a la que puede producirse el entrecruzamiento se llama frecuencia recombinante.
Para que se pueda notar el hecho de la recombinación, es necesario que haya en
el cromosoma por lo menos dos loci en heterocigosis.
Por el proceso de crossing-over, genes que están ligados pueden dejar de
estarlo y recombinarse con otros genes, logrando así que los caracteres que
rigen se transmitan de manera independiente, es decir, que se cumpla en ellos
la 3ª ley de Mendel. En los cromosomas que no ocurra entrecruzamiento, cada una
de sus cromátidas formará un cromosoma "puro" en el gameto al que
vaya a parar, mientras que las cromátidas no hermanas intercambiadas formarán
un cromosoma "mixto", es decir, con intercambio de genes.
El mapa cromosómico o genético consiste en la representación gráfica de
los diferentes cromosomas de esa especie, indicando en cada cromosoma los genes
que están presentes, sus correspondientes alelos, su localización, orden y
distancia relativa entre unos y otros genes (distancia génica).

Durante el proceso de ligamiento en la meiosis pueden aparecer dos clases de
progenie: Parentales, con la misma combinación de alelos que sus padres (con
gametos no producidos por recombinación) y recombinantes, con diferente
combinación de alelos que sus padres por entrecruzamiento (con gametos
producidos por recombinación).

La posibilidad de intercambio génico aumenta con la distancia en el cromosoma
entre esos genes. La distancia génica se mide en centimorgan (cM). El tamaño
medio de un cromosoma es de 50 Mb (megabases). Físicamente, 1 cM corresponde a
una secuencia de ADN de entre 0.7 y 1.2 megabases.
Se dice que dos “loci” están a un cM de distancia si su frecuencia de
recombinación es una de cada 100 meiosis. Por cada centimorgan, el porcentaje
de recombinación aumenta en un 1%, es decir, que un 1% de fenotipos recombinantes
se corresponde con un centimorgan. La distancia génica máxima que se considera
entre dos genes es de 50 centimorgan (tamaño medio del cromosoma) por lo que,
en ningún caso, los dos nuevos fenotipos surgidos del “crossing over” superarán
en porcentaje a los parentales que no han sufrido dicho “crossing over”. Como
máximo, presentarán el mismo porcentaje (25% para cada uno de los dos fenotipos
recombinantes).
Calculando el porcentaje de recombinación entre los diversos genes, se puede
construir el mapa cromosómico o genético de esa especie. En un futuro no muy
lejano será posible establecer el mapa génico del canario con todo detalle. Así
se podrá mejorar la raza de forma controlada e, incluso, mediante la ingeniería
genética, propiciar la aparición de nuevas mutaciones y caracteres, o prevenir
enfermedades y malformaciones genéticas.
La representación esquemática de los
genes y fórmulas...
En genética teórica y aplicada es costumbre utilizar fórmulas y esquemas en los
que los genes estudiados son representados por letras o símbolos derivados
generalmente del nombre de los caracteres fenotípicos afectados en cada caso.
La notación en letra mayúscula representa su carácter dominante, siendo en
minúscula el recesivo. El exponente “+” representa gen silvestre no mutado. El
término “portador” se representa por el símbolo “/”.
Los caracteres que afectan al lipocromo son:

Los caracteres que afectan a las melaninas son:

Según la nomenclatura anterior el canario original silvestre macho tendría la
siguiente fórmula genética:
E+ (Z) z+
rb+ G+ B+ I+
--------------------------------------
E+ (Z) z+ rb+ G+ B+ I+
Dónde:
E+ = Presencia de eumelaninas.
(Z) = Cromosoma sexual Z.
z+ = Eumelanina negra (no mutado).
rb+ = Oxidación melánica (no mutado).
G+ = Lipocromo amarillo.
B+ = Presencia de feomelanina.
I+ = Nevado.
Al ser homocigótico, los dos elementos del quebrado (que representan ambos
alelos) son iguales.
Teniendo en cuenta las leyes de Mendel, para calcular el resultado de un cruce
determinado, se utilizan las tablas gaméticas de Punnett, trabajando con los
genes que pueden sufrir modificación en el cruce, ya que el resto quedarían
inalterados.
El cuadro de Punnett es un diagrama diseñado por Reginald Punnett, usado por
los biólogos para determinar la probabilidad dominante, codominante o recesiva.
Por ejemplo, en el cruce de dos canarios píos, cuyo genotipo sería:
E+
-----
E
Dónde:
E+ = Presencia de melaninas.
E = Ausencia de melaninas (mutado).
Si se cruzan estos píos, según la 2ª ley de Mendel, en la tabla de Punnett se
representaría así:

La probabilidad de que el producto tenga el genotipo E+E+ (melánico puro) es de
25%, con E+E (pío) es del 50% y con EE (lipocromo puro) es del 25%. Todos los
genotipos son alelos, por lo tanto todos son conocidos como un Punnett normal o
adyacente.
Poniendo otro ejemplo, un poco más complejo, se va a utilizar un macho de
fenotipo ágata amarillo intenso (genotipo to+ to+ I I) con una hembra de
fenotipo ágata topacio amarillo nevado (genotipo to to I+ I+). Todos los
pájaros resultantes tendrán el gen “ágata” y el amarillo en su genotipo, por lo
que se han cogido los genes topacio mutado y no mutado), intenso y nevado que
son los que sufrirán modificaciones. El resultado de este cruce serán pájaros F1
cuyo fenotipo será para el 100% de los ejemplares ágata amarillo
intenso/topacio y nevado (el símbolo”/” significa portador, por tanto serán
portadores de topacio y de nevado, con genotipo to+ to I+ I). Estos
pájaros F1 resultantes son ejemplares dihíbridos, es decir, heterocigóticos
para dos genes distintos (to+ e I+).
Si volvemos a cruzar dos F1, ambos con ese mismo genotipo (to+ to I+
I), aplicaría la 3ª ley de Mendel. La tabla de Punnett quedaría como
sigue:

Ya que los alelos dominantes eclipsan a los recesivos (habría que analizar
posteriormente los que son portadores y de qué gen) hay nueve combinaciones que
tienen el fenotipo ágata amarillo intenso (los de color verde en la tabla),
tres que son ágata amarillo nevado (los de color azul en la tabla), tres ágata
topacio amarillo intenso (los de color rojo en la tabla) y uno ágata topacio
amarillo nevado (el de color negro en la tabla). La proporción se muestra como
9:3:3:1 y es la más usual para el cruce dihíbrido, siempre que no ocurra epistasia,
como se explicó anteriormente.
Tabla que resume los caracteres hereditarios que afectan al canario de color y
su transmisión genética:

De ella podéis deducir cuándo un carácter va a quedar
"oculto" porque su gen recesivo se ha unido a otro dominante, y
cuándo existe codominancia (saliendo canarios naranjas, píos, etc...).
Articulo subido por Weasel en http://foro-cria-de-canario.foroactivo.com